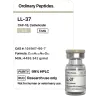

LL-37
For in vitro testing and laboratory use only. Not for human or animal consumption. Bodily introduction is illegal. Handle only by licensed professionals. Not a drug, food, or cosmetic. Educational use only.
LL-37: Small Peptide, Big Research Interest
LL-37 is not just another "hyped" peptide, but the body's own human cathelicidin: a small yet extremely busy molecule of innate immunity. In experimental models and preclinical data, it has shown activity across several areas at once — from antimicrobial effects and anti-biofilm applications to involvement in inflammation, angiogenesis, re-epithelialization, and tissue repair processes.
But the real intrigue is that LL-37 does not follow simple rules: in a research context, its effects depend on the environment, the tissue, and the specific model. Early clinical observations in hard-to-heal venous ulcers produced an encouraging signal, although a larger study did not confirm a statistically significant advantage across the full group.
This is precisely why LL-37 stands out in scientific discussion: not because it offers an easy narrative, but because it combines impressive biological breadth with a high degree of context dependence. It is a molecule that has already demonstrated striking biology in the research setting, yet still requires a careful and intelligent approach.
Practical Takeaway
For the client, this is exactly the kind of case where interest is driven not by marketing noise, but by genuinely strong research potential.
LL-37 (Human Cathelicidin / hCAP18): A Scientific Review
Based on peer-reviewed literature — see References. Last updated: April 2026.
The Short Version
LL-37 is the human body’s only cathelicidin — the sole representative of an ancient family of antimicrobial peptides. It is not a synthetic pharmaceutical candidate designed in a laboratory. It is an endogenous effector of the innate immune system, produced by neutrophils, epithelial cells, and multiple other tissues.
What makes LL-37 simultaneously fascinating and difficult to develop therapeutically is a paradox that runs through its entire biology: the same molecule that protects is also pathological. At the right concentration, in the right location, LL-37 kills bacteria, disrupts biofilms, accelerates wound healing, and modulates immune responses appropriately. At the wrong concentration or location, it drives the chronic inflammation of rosacea, amplifies psoriasis flares through plasmacytoid dendritic cell activation, contributes to atherosclerosis by promoting LDL uptake, and may promote tumour growth in some cancers while killing tumour cells in others.
Three biological threads define LL-37’s importance: extraordinary antimicrobial breadth (effectively combating over 38 bacteria, 16 fungi, and 16 viruses through membrane disruption and biofilm suppression[11]); direct regulation by vitamin D through the CAMP gene promoter (providing a mechanistic explanation for the epidemiological associations between vitamin D deficiency and infection susceptibility[5]); and active disease contribution when dysregulated (in rosacea, psoriasis, IBD, RA, and atherosclerosis[8]).
| At a glance | |
|---|---|
| Common names | LL-37; human cathelicidin; hCAP18 (precursor) |
| Gene | CAMP (Cathelicidin Antimicrobial Peptide); chromosome 3p21.3 |
| Precursor protein | hCAP18 (18 kDa; 170 amino acids) |
| Active peptide | LL-37 (4.5 kDa; 37 amino acids) |
| Cleavage enzyme | Proteinase 3 (neutrophils); kallikrein 5/7 (skin keratinocytes) |
| Sequence | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES |
| Structure | Amphipathic α-helix; cationic; unstructured in aqueous solution, helical at membranes |
| Key regulator | Vitamin D (1,25-dihydroxyvitamin D&sub3; → VDR → CAMP promoter) |
| FDA status | ❌ Not approved for any indication |
| Clinical trial status | Phase 1 completed (melanoma, intratumoural); no therapeutic approvals |
Biology: The Only Human Cathelicidin
The cathelicidin family
Cathelicidins are an evolutionarily ancient family of antimicrobial peptides found across vertebrates. Most mammals possess multiple cathelicidins — pigs have over 10, cattle have several, mice have one (CRAMP). Humans have precisely one: the product of the single CAMP gene on chromosome 3p21.3, encoding hCAP18 (170 amino acids). hCAP18 is a major component of the azurophilic granules of neutrophils and is primarily produced by bone marrow, keratinocytes of inflamed sites, and cells of the mucosal epithelium.[1] The conserved N-terminal “cathelin domain” acts as an inactive protective prodomain; all biological activity resides in the variable C-terminal 37-amino acid fragment released by proteolytic cleavage.
From gene to active peptide: the processing cascade
CAMP gene transcription is induced primarily by vitamin D (via VDR), but also by innate immune signals, microbial products, butyrate (from gut bacteria), and cytokines. hCAP18 is synthesised as a preproprotein, packaged into neutrophil secondary granules, and stored inactive. Upon degranulation, hCAP18 is secreted: in neutrophils, proteinase 3 cleaves the cathelin domain, releasing active LL-37. In skin keratinocytes, kallikreins 5 and 7 perform this cleavage — a tissue-specific protease difference that partly explains why skin-derived LL-37 processing is dysregulated in rosacea (overactive KLK5) vs. the neutrophil system.
Structure and amphipathicity: why it kills membranes
In aqueous solution, LL-37 is largely disordered. Upon encountering a membrane, it adopts an amphipathic α-helical conformation with positively charged residues on one face and hydrophobic residues on the other. The peptide’s selectivity exploits the fundamental difference between bacterial membranes (anionic — attracting the cationic +6 charge LL-37) and host cell membranes (predominantly neutral outer leaflet). At physiological concentrations this selectivity provides a therapeutic window — though at high concentrations LL-37 is cytotoxic to host cells as well.
Mechanisms of Action
Antimicrobial — membrane disruption
LL-37 disrupts bacterial membranes through the carpet mechanism (coating membrane surface in a detergent-like manner until solubilised), toroidal pore formation (transient pores allowing ion efflux and membrane potential collapse), and membrane thinning. Critically, these mechanisms target membrane structure rather than specific molecular targets — which is why LL-37 retains activity against antibiotic-resistant organisms where resistance mechanisms based on modifying a specific enzyme or effluxing antibiotic molecules are irrelevant.[4]
Anti-biofilm
LL-37’s anti-biofilm activity at concentrations well below those required for direct bactericidal activity is one of its most clinically relevant properties. LL-37 has been shown to have antimicrobial and anti-biofilm activity against multiple Gram-positive and Gram-negative human pathogens, and can eradicate preformed biofilms in vitro.[4] The mechanism involves interference with quorum sensing, disruption of extracellular polysaccharide matrices, and inhibition of biofilm attachment to surfaces — making it potentially valuable for chronic infected wounds where biofilm formation is the primary reason antibiotic treatment fails.
Immunomodulatory — the inflammatory switch
LL-37 is a multifunctional immunomodulator connecting innate and adaptive immunity. It is a chemoattractant for neutrophils, monocytes, mast cells, and T cells. It modulates Toll-like receptor signalling in a context-dependent manner — it can suppress LPS-induced TLR4 activation (anti-inflammatory in endotoxaemia) or amplify TLR9 and TLR7 signalling when complexed with self-DNA (pro-inflammatory in psoriasis and lupus).[9] When LL-37 binds extracellular DNA from damaged cells, the LL-37/DNA complex enters plasmacytoid dendritic cells (pDCs) and activates TLR9, driving type I interferon production — a mechanism central to psoriasis pathogenesis.
Wound healing
High levels of hCAP18 are produced in skin upon wounding, with the highest levels at 48 hours post-injury, declining to pre-injury levels upon wound closure. The immunoreactivity for hCAP18/LL-37 is absent in chronic ulcer edge epithelium.[3] Wound healing effects involve direct stimulation of keratinocyte migration and proliferation via EGFR/ErbB transactivation; VEGF-A upregulation (neovascularisation); TGF-β upregulation (granulation tissue); and direct antimicrobial protection preventing wound colonisation from impeding healing. The observation that chronic ulcers lack LL-37 expression provides mechanistic rationale for topical LL-37 in non-healing wounds.
The Vitamin D–LL-37 Axis
The CAMP gene promoter contains functional vitamin D response elements (VDREs), and binding of the active metabolite 1,25(OH)&sub2;D&sub3; to the vitamin D receptor (VDR) directly drives CAMP gene transcription.[5] Plasma hCAP18 concentrations correlate with serum 25-hydroxyvitamin D concentrations in subjects with 25(OH)D levels ≤ 32 ng/mL (r = 0.81, p < 0.005) but not in subjects above this level — consistent with a receptor-mediated response with a ceiling effect once VDR is fully occupied.[6] In dialysis patients with very low vitamin D, those with the lowest hCAP18/LL-37 levels had a greater than 2-fold increased risk of death from infectious causes.
This vitamin D/LL-37 relationship provides a mechanistic explanation for vitamin D’s role in infection susceptibility that goes beyond correlational epidemiology: vitamin D deficiency reduces innate antimicrobial capacity at the level of cathelicidin production. It also provides a plausible mechanism for the historical observations that sun exposure (UVB → vitamin D synthesis → LL-37 upregulation) reduces tuberculosis severity and may protect against respiratory infections.
LL-37 in Disease: When the Protector Becomes the Problem
Rosacea: dysregulated processing, not excess production
Rosacea is mechanistically one of the best-characterised LL-37 diseases. Environmental factors activate TLR2 on keratinocytes, leading to kallikrein 5 expression. KLK5 cleaves hCAP18, producing LL-37, triggering the NLRP3 inflammasome, JAK/STAT, and NF-κB pathways. LL-37 stimulates mast cells via MRGPRX2, inducing inflammatory cytokines and MMP-9. MMP-9 further enhances KLK5 production, creating a positive feedback loop.[7] The pathological LL-37 in rosacea differs from normal wound-healing LL-37 in quantity, processing protease, and location — the KLK5-generated fragment (“FF/LL-37”) is a more potent inducer of mast cell degranulation and vascular dilation than canonical LL-37, explaining the erythema and telangiectasias of rosacea. This mechanistic understanding explains why ivermectin (reducing Demodex density) and tetracyclines (inhibiting MMPs) are effective.
Psoriasis: LL-37 as the autoimmune trigger
In psoriasis, LL-37 is overproduced in keratinocytes and forms complexes with self-DNA released from damaged cells. These LL-37/DNA complexes enter pDCs via TLR9, triggering type I interferon production and breaking tolerance to self-DNA; activate TLR2, TLR4, and inflammasome pathways in keratinocytes; and stimulate Th17 and Th1 responses. LL-37 is therefore both a consequence of psoriatic inflammation (produced by inflamed keratinocytes) and a cause of the autoimmune amplification (activating pDCs to break self-tolerance). Serum hCAP18/LL-37 levels are significantly elevated in psoriasis patients compared to healthy controls across multiple studies.
Cardiovascular risk: LL-37 and atherosclerosis
Serum hCAP18/LL-37 levels are significantly higher in patients with psoriasis, rosacea, IBD, and RA than in healthy individuals. LL-37 positively correlated with PC-oxidised phospholipid (PC-OxPL) levels, a predictive factor for the development and progression of atherosclerosis.[8] The proposed mechanism: LL-37 binds to LDL particles, causing conformational changes that enhance LDL uptake into macrophages and endothelial cells beyond normal LDL receptor pathways, driving foam cell formation — the earliest step in atherosclerotic plaque development. This suggests that the elevated cardiovascular risk observed in psoriasis patients may be mechanistically mediated, at least in part, by chronically elevated LL-37.
Cancer: profoundly context-dependent
LL-37’s role in cancer is bidirectional and cancer-type specific. In pro-cancer contexts, LL-37 promotes tumour cell proliferation and angiogenesis via ErbB and EGFR signalling in ovarian cancer, breast cancer, and lung cancer cell lines. In anti-cancer contexts, LL-37 directly lyses tumour cells via membrane disruption, activates pDCs, stimulates NK cell activity, and expands antigen-specific CD8+ T cells. A Phase 1 clinical trial explored intratumoural LL-37 injection in melanoma patients — the only completed human therapeutic trial — based on this immunostimulatory rationale.[10]
Clinical Evidence: What Has Been Tested in Humans
Phase 1 melanoma trial
The only published human interventional trial of therapeutic LL-37 used intratumoural injection in melanoma patients with cutaneous metastases. The trial was Phase 1 (safety/dose-finding) and demonstrated safety and tolerability at doses tested, with immunological evidence of anti-tumour immune activation. No Phase 2 follow-up has been published.
Clinical significance from deficiency and excess states (observational)
The strongest human evidence for LL-37’s biological importance comes from observational data at the extremes of expression. Morbus Kostmann (severe congenital neutropenia): Patients have functionally absent LL-37 in neutrophils and suffer severe recurrent periodontal disease disproportionate to their neutrophil counts — confirming LL-37 is not redundant but essential. Atopic dermatitis: LL-37 is deficient in atopic skin, explaining dramatically increased susceptibility to skin infections (particularly S. aureus and eczema herpeticum from HSV); AD patients have 30-fold lower cathelicidin expression in acute lesions compared to psoriasis — explaining why psoriasis is rarely complicated by secondary infection while AD frequently is. Chronic wounds: LL-37 is absent in chronic ulcer epithelium while present at high levels in normal wound healing, providing human in vivo evidence for its role in re-epithelialisation.[3]
Evidence Summary
| Evidence type | Finding | Quality |
|---|---|---|
| In vitro antimicrobial [11] | Kills >38 bacteria, 16 fungi, 16 viruses via membrane disruption | Strong; extensively replicated |
| In vitro anti-biofilm [4] | Disrupts preformed biofilms at sub-MIC concentrations | Strong; multiple labs |
| Human wound healing (observational) [3] | hCAP18 absent in chronic ulcers; present in normal healing | Strong observational |
| AD deficiency (human) | Deficient cathelicidin correlates with infection susceptibility | Strong observational |
| Rosacea pathogenesis [7] | LL-37 fragments drive rosacea inflammation (mechanistic + human correlative) | Strong |
| Psoriasis & atherosclerosis (human) [8] | Elevated serum hCAP18; LL-37 promotes LDL uptake; correlates with PC-OxPL | Strong observational + mechanistic |
| Vitamin D correlation (human) [6] | Circulating hCAP18 correlates with 25(OH)D ≤32 ng/mL | Moderate (multiple replications) |
| Cancer (Phase 1) [10] | Intratumoural LL-37 safe in melanoma; immunological activation | Phase 1 only; no efficacy data |
LL-37 Analogues: The Clinical Development Path
The clinical application of native LL-37 is hindered by low proteolytic stability, cytotoxicity, and high production costs. Multiple design strategies are actively being explored. Truncated peptides (LL-17, GF-17, FK-16, LL-23) retain antimicrobial or wound-healing activity with reduced cytotoxicity. All-D enantiomers (retro-inverso peptides) resist host proteases due to their stereospecificity, dramatically improving biological stability. Stapled peptides constrain the bioactive α-helical conformation; a glycosylated double-stapled analogue (SLP-51) showed 35-fold enhanced antimicrobial potency compared to the linear parent with minimal haemolysis in 2025 preclinical work.[9] Nanotechnology delivery (liposomes, chitosan-based hydrogels) achieves localised controlled release in wounds, allowing high local concentrations without systemic exposure.
Common Misconceptions
“LL-37 is an antibiotic.”
LL-37 is an antimicrobial peptide, mechanistically distinct from antibiotics. Antibiotics typically target specific bacterial proteins; LL-37 targets membrane structure. The consequence is that LL-37 retains activity against MRSA, carbapenem-resistant organisms, and other multidrug-resistant bacteria that have rendered conventional antibiotics ineffective.
“Higher LL-37 is always better.”
⚠️ Emphatically false. The diseases in which LL-37 is pathologically elevated — rosacea, psoriasis, rheumatoid arthritis, IBD, and potentially atherosclerosis — demonstrate that excess LL-37 drives chronic inflammation and cardiovascular disease. Physiological LL-37 is tightly regulated precisely because both deficiency and excess produce pathology.[8]
“Taking LL-37 supplements will boost your immune system.”
⚠️ There is no evidence that oral LL-37 supplementation produces systemic immunological effects. LL-37 is a peptide and would be proteolytically degraded in the gastrointestinal tract. There is no established oral bioavailability. The wellness market sale of LL-37 as an oral supplement has no clinical evidence basis.
“LL-37 is safe because it’s natural.”
At physiological concentrations in appropriate locations, LL-37 is well-tolerated. At supraphysiological concentrations, it is cytotoxic to osteoblasts, haemolytic toward red blood cells, and inflammatory in tissues where it should not be present at high levels. The cytotoxicity is concentration- and context-dependent — not absent.
Frequently Asked Questions
Why doesn’t LL-37 kill human cells when it kills bacteria?
The relative selectivity at lower concentrations arises from the difference in membrane composition: bacterial membranes are predominantly anionic, attracting the cationic LL-37 more strongly. Eukaryotic cell membranes are predominantly zwitterionic on the extracellular face, producing less electrostatic attraction. This creates a concentration window where bacteria are killed and host cells are not — a window that narrows as concentration rises.
Can vitamin D supplementation meaningfully increase LL-37 levels?
In vitamin D deficient individuals (25(OH)D ≤ 32 ng/mL), supplementation that raises vitamin D levels does appear to increase circulating hCAP18/LL-37.[6] In already-sufficient individuals, the correlation saturates and supplementation produces minimal additional LL-37 effect. Whether this LL-37 increase meaningfully reduces infection susceptibility has been studied most extensively in the context of tuberculosis, with some positive findings in deficient patients, but is not definitively established.
Is LL-37 relevant to MRSA treatment?
LL-37 retains antimicrobial activity against MRSA in vitro. However, the MIC concentrations required are at or above concentrations producing cytotoxicity in host cells, and S. aureus has multiple LL-37 resistance mechanisms (including D-alanylation of teichoic acids, protease secretion, and carotenoid production). The anti-biofilm properties at sub-MIC concentrations may be the most clinically relevant anti-MRSA application.[4]
Key Takeaways
- LL-37 is the sole human cathelicidin — a 37-amino acid endogenous antimicrobial peptide with an amphipathic α-helical structure enabling membrane-disrupting activity against bacteria, fungi, viruses, and biofilms. Its unique role as the only human representative of an otherwise diverse family makes its biology particularly important.[1]
- The vitamin D–LL-37 axis is a physiologically important regulatory circuit. Vitamin D directly drives CAMP gene transcription. Vitamin D deficiency impairs innate antimicrobial capacity at the level of cathelicidin production — providing a mechanistic explanation for vitamin D’s role in infection susceptibility.[5]
- ⚠️ LL-37 is pathological when dysregulated. In rosacea (abnormal KLK5 processing), psoriasis (LL-37/DNA complex activating pDCs), and atherosclerosis (LL-37-mediated LDL uptake), LL-37 is not merely a bystander but an active driver of disease. Exogenous LL-37 could worsen rosacea and psoriasis, and elevated levels contribute to cardiovascular risk.[7][8]
- The clinical evidence is primarily observational and preclinical. The human evidence for LL-37’s biological importance comes from deficiency states (Kostmann, AD, chronic wounds) and excess states (psoriasis, rosacea), not from controlled therapeutic trials. The only completed human therapeutic trial is a Phase 1 intratumoural melanoma study.
- The path to clinical development runs through analogues and delivery systems, not native LL-37. The obstacles — context-dependence, selectivity window, production cost, protease instability — are real. Modified analogues with improved stability, reduced cytotoxicity, and lower cost are the active research frontier.[9]
- ⚠️ Community use of injectable LL-37 has no validated evidence base — no pharmacokinetic data in humans, no dose-response characterisation outside Phase 1 oncology, and an acknowledged cytotoxicity profile that makes systemic use at therapeutic concentrations potentially harmful.
References
Foundational Biology
- Frohm M, Agerberth B, Ahangari G, et al. The expression of the gene coding for the antibacterial peptide LL-37 is induced in human keratinocytes during inflammatory disorders. Journal of Biological Chemistry. 1997;272(24):15258–15263.
- Sørensen OE, Cowland JB, Theilgaard-Mönch K, et al. Wound healing and expression of antimicrobial peptides/polypeptides in human keratinocytes, a consequence of common growth factors. Journal of Immunology. 2003;170(11):5583–5589.
Wound Healing
- Sørensen OE, Thapa DR, Rosenthal A, et al. Differential regulation of cathelicidin LL-37 in human skin. Journal of Investigative Dermatology. 2003. PMID: 12603850
Anti-Biofilm and Wound Infection
- Dean SN, Bishop BM, van Hoek ML. The human cathelicidin antimicrobial peptide LL-37 as a potential treatment for polymicrobial infected wounds. Frontiers in Immunology. 2013. PMC3699762
Vitamin D Regulation
- Gombart AF, Borregaard N, Koeffler HP. Human cathelicidin antimicrobial peptide (CAMP) gene is a direct target of the vitamin D receptor and is strongly up-regulated in myeloid cells by 1,25-dihydroxyvitamin D&sub3;. FASEB Journal. 2005;19(9):1067–1077.
- Positive correlation between circulating cathelicidin antimicrobial peptide (hCAP18/LL-37) and 25-hydroxyvitamin D levels in healthy adults. BMC Research Notes. 2012. PMC3532295
Disease States
- Therapeutic strategies focusing on immune dysregulation and neuroinflammation in rosacea. PMC11317294. 2024.
- Increased LL37 in psoriasis and other inflammatory disorders promotes LDL uptake and atherosclerosis. Journal of Clinical Investigation. 2024.
Comprehensive Reviews
- Voronko OE, et al. Antimicrobial Peptides of the Cathelicidin Family: Focus on LL-37 and Its Modifications. International Journal of Molecular Sciences. 2025;26(16):8103. PMC12386566
- Renovation as innovation: Repurposing human antibacterial peptide LL-37 for cancer therapy. Frontiers in Pharmacology. 2022. PMC9445486
- Decoding LL-37 review — LL-37 effectively combats over 38 bacteria, 16 fungi, and 16 viruses. ScienceDirect. 2025.
Certificate of Analysis
An independent test report is available for LL-37 5mg. This report provides batch-level documentation and analytical verification information for research reference.

Based on 1 reviews
5.0
The cationic amphipathic helix is the architecture we actually study — LL-37 folds into an α-helix on membrane contact, the cationic face engaging the negatively-charged bacterial membrane and the hydrophobic face inserting into the lipid bilayer. For probing membrane-disruption mechanism across antimicrobial-peptide structural classes, LL-37 is the human-cathelicidin reference because it's the only one humans have. The material delivered the expected membrane-permeabilization in our bacterial-membrane model assays.
Yes. It is a 37-amino-acid human host-defense peptide, the active part of the cathelicidin hCAP18/CAMP.
No, as a standalone drug LL-37 is not approved by the FDA or EMA.
In laboratory and preclinical models — yes, and not only planktonic bacteria, but in some cases biofilms as well. But that is not equivalent to proven clinical efficacy as an antibiotic in humans.
Potentially yes, and the early clinical signals were encouraging. But a larger study did not confirm a significant benefit in the overall population of patients with venous ulcers.
Yes, as a research hypothesis and early translational direction — yes. But LL-37 may also have protumorigenic effects in certain tissues and models. This is very much not a topic where simple conclusions are appropriate.
LL-37 is a biologically very important human molecule and an interesting research candidate, but at present there are not enough data to view it as a proven universal therapy in humans
LL‑37 is processed from hCAP18, the only cathelicidin found in humans, and is expressed by immune cells such as neutrophils and epithelial cells. It plays an important role in the innate immune defense against infection by interacting with microbial membranes.
LL‑37 displays broad antimicrobial activity against many bacteria and other pathogens. It also interacts with host cells to modulate immune responses and may influence processes like chemotaxis, inflammation, and wound healing.
In research, LL‑37 is usually supplied as a lyophilized (freeze‑dried) peptide powder.
Commercial peptide listings typically report LL‑37 purity at ≥ 96 % (e.g., by UHPLC) for research applications, though exact values may vary by supplier and batch.
As a research peptide product, LL‑37 is usually labeled for laboratory or research use only and not for human or veterinary therapeutic use unless otherwise specified by regulatory approval. (This is standard for research‑grade peptides — check your product label/policy.)
LL-37 is a 37-amino acid cationic antimicrobial peptide (AMP) and the sole member of the human cathelicidin family. It is generated in vivo by extracellular proteolytic cleavage of the C-terminal domain of a larger precursor protein called hCAP18 — primarily by serine proteases of the kallikrein family in keratinocytes and by proteinase 3 in neutrophils. It is produced by multiple cell types including neutrophils, macrophages, mast cells, natural killer cells, and epithelial cells throughout the respiratory, gastrointestinal, and urogenital tracts, placing it at the front line of innate immune defense. It was first characterized in 1995 and has since become one of the most extensively studied antimicrobial peptides in biomedical research, with over 1,800 published studies. Despite this research depth it has not yet achieved regulatory approval as a therapeutic agent for any indication.
LL-37 operates through multiple parallel mechanisms that distinguish it from most conventional antibiotics. Its primary antimicrobial mechanism relies on its amphipathic alpha-helical structure and net positive charge of +6, which allow it to electrostatically bind the negatively charged membranes of bacteria, fungi, and enveloped viruses — cell types whose membranes differ fundamentally from the cholesterol-rich membranes of human cells. Upon binding it inserts into the microbial membrane, forming transmembrane pores that disrupt membrane integrity, cause cellular leakage, and ultimately lyse the microorganism. It can also penetrate bacterial cells to interact with intracellular targets including nucleic acids. Against biofilms — structured bacterial communities that resist conventional antibiotics — it disrupts the extracellular matrix that holds the biofilm together, exposing the underlying bacteria to immune attack and antibiotics. Beyond direct killing, LL-37 activates the FPRL-1 receptor (a G protein-coupled receptor) to recruit neutrophils, monocytes, and T cells to infection sites. It stimulates the P2X7 receptor to promote IL-1β secretion. It activates epidermal growth factor receptor and ERK/MAPK signaling to promote wound healing and epithelial repair. It binds IGF-1 receptor to drive further cell survival and proliferation signaling.
A 2025 systematic review through ScienceDirect documented LL-37's antimicrobial activity against over 38 bacterial species, 16 fungal species, and 16 viral species. Bacteria against which activity has been documented include Staphylococcus aureus, Staphylococcus epidermidis, Escherichia coli, Pseudomonas aeruginosa, Enterococcus, and multiple antibiotic-resistant strains. Antiviral activity has been documented against influenza A, herpes simplex viruses, HIV, and respiratory syncytial virus — primarily through disruption of viral envelopes and interference with post-entry replication steps. Antifungal activity has been shown against Candida species. Particularly noteworthy is its anti-biofilm activity, which is clinically relevant for chronic wound infections, device-associated infections, and pulmonary infections in cystic fibrosis where conventional antibiotics fail against biofilm-protected organisms.
LL-37's immunomodulatory properties have generated research interest in wound healing — it promotes angiogenesis, stimulates keratinocyte migration and proliferation, and accelerates re-epithelialization of chronic wounds including diabetic ulcers. Cancer research has explored both its tumor-suppressive effects in some contexts and its tumor-promoting effects in others — the paradox of a host defense peptide whose pro-inflammatory and angiogenic properties can, depending on the cancer type and microenvironment, either suppress or promote malignancy. Autoimmune research has documented LL-37's role as a driver of lupus, psoriasis, and rheumatoid arthritis pathogenesis — in psoriasis elevated LL-37 forms complexes with self-DNA that activate plasmacytoid dendritic cells through TLR9, driving chronic type I interferon production and inflammation. This makes LL-37 simultaneously a defender against pathogens and a potential participant in autoimmune disease pathogenesis.
Four challenges have consistently blocked LL-37 from reaching approved therapeutic status. Production cost is high — synthesizing a 37-amino acid cationic peptide at pharmaceutical scale is expensive. Proteolytic instability in physiological environments — particularly in the presence of host serine proteases and in the high-salt conditions of serum — significantly reduces its activity and half-life in vivo. Cytotoxicity to human cells at higher concentrations is a dose-limiting concern, as the same membrane-disrupting mechanism that kills bacteria can also affect mammalian cells at elevated concentrations. Reduced efficacy in the physiological ionic strength of human serum and tissue fluids compared to laboratory buffers is a consistent finding. Active research into LL-37 analogs, truncated derivatives, lipid nanoparticle delivery systems, and chemical modifications aims to address these limitations.
At therapeutic research doses LL-37 generally shows a favorable side effect profile. The most commonly reported effects are mild injection site reactions including redness, swelling, and discomfort. Headaches have been reported. Mild allergic reactions are possible given its immunomodulatory activity. The primary safety concern is cytotoxicity — at higher concentrations LL-37 can damage human cell membranes through the same mechanism it uses against bacteria, making dose precision important. The autoimmune concern is also relevant in therapeutic contexts — in individuals with predisposition to lupus, psoriasis, or other autoimmune conditions, exogenous LL-37 supplementation could theoretically amplify the self-DNA sensing and type I interferon pathways that drive these conditions.
People with active autoimmune conditions — particularly systemic lupus erythematosus, psoriasis, or rheumatoid arthritis — should not use it given LL-37's documented role in amplifying the innate immune pathways that drive these diseases. People with active cancer should exercise significant caution given the complex and context-dependent cancer biology — LL-37 has been shown to promote tumor progression in some cancer types including lung and ovarian cancer through its angiogenic and EGFR/IGF-1R signaling properties. Pregnant or breastfeeding women should not use it given insufficient safety data. It is not FDA-approved for any therapeutic indication and should only be used within formally approved research settings.